
��(d��ng)ǰλ�� > ���(y��) > ���g(sh��)���� > ����FGFR3����^(q��)�����scFv���w�ĺY�x�ͻ����о�
����FGFR3����^(q��)�����scFv���w�ĺY�x�ͻ����о�
����FGFR3����^(q��)�����scFv���w�ĺY�x�ͻ����о�
-Molecular Devices
���w�S��(x��)�����L(zh��ng)����(FGF)������һ����ߌӺ���Ƥ��(x��)�����L(zh��ng)�ֻ��������Ӽ��塣FGFs���S������W(xu��)�^���аl(f��)�]��Ҫ���ã�������̥�l(f��)��������������Ѫ��(x��)�����ɼ�Ѫ�ܰl(f��)�����������������о�����FGFs�������Ӷ�N�[����(x��)���Ľ���(r��n)������ǰ�������������I�K���鷿�����ٵȲ�λ�[����
Ŀǰ�����l(f��)�F(xi��n)20��NFGFs����(du��)��ͬ��ͼ�(x��)�����в�ͬ��������������ֻ�l(f��)�F(xi��n)��5�NFGF���w(FGFR)���ڵ���ˮƽ�����@Щ���w����55%-72%��ͬԴ�ԡ�FGFR�Y(ji��)��(g��u)����һ��(g��)�������w�Y(ji��)�υ^(q��)����һ��(g��)��Ĥ�^(q��)���Լ�һ��(g��)����(n��i)��ø�^(q��)�������w�Y(ji��)�υ^(q��)���������(g��)��ͬ����������Y(ji��)��(g��u)��(�Q��������I��II��III)��FGFR1-3 mRNA�IJ�ͬ�ļ��������γɃɷN���ͦ��ͦ�������FGFR3���ЃɷN��ͬ��ͻ׃�wIIIb��IIIc���@�ɷN׃�w���в�ͬ���H�ͻ��ԣ�IIIc�ֲ����ӏV�����ܺͶ�NFGFs�Y(ji��)��(FGF1��F(xi��n)GF2��F(xi��n)GF4����FGF9)��IIIb��(y��u)�Ⱥ�FGF1�Y(ji��)�����܉��^�ͳ̶Ⱥ�FGF8��FGF9�Y(ji��)�ϡ����и���(heparin)���õ���r����F(xi��n)GFs��FGFRs�Y(ji��)�Ϻ���F(xi��n)GFs�T��(d��o)���w���ۻ����������(n��i)��ø�^(q��)����������ữ�Լ�������̖(h��o)��(j��)(li��n)����(y��ng)�ļ��������w���w�Y(ji��)�Ϻ�F(xi��n)GFs��(hu��)����(d��ng)��N��̖(h��o)�D(zhu��n)��(d��o);��������(n��i)�}�x��ˮƽ�������T��(d��o)�z��ԭ�����ø�͵���øCͨ·������������h(hu��n)��ø�Լ��T��(d��o)ԭ������c-myc ��c- fos��
�Ѱl(f��)�F(xi��n)FGFR3��(hu��)�l(f��)������ͻ׃����(d��o)�����Ұ��ἤø���Լ������һЩ�c�����l(f��)������l(f��)�Թ��������i���[���Լ������[�����P(gu��n)�ľC������������о��l(f��)�F(xi��n)FGFR��̖(h��o)��ǰ�����[����(x��)�����w��������ȫ�������������F(xi��n)GFR3�ѱ������l(f��)�Թ������Ŀ����ί����c(di��n)���M�����д_��(sh��)�C��(j��)���������FGFR3ͻ׃�������[���M�����������P(gu��n)��FGFR3���[���M���еı��_(d��)֪���ķdz��١�������ʹ�û���оƬ���g(sh��)��(du��)������_(d��)���������l(f��)�F(xi��n)�������M�����FGFR3�ڰ����[����Ʒ���^���_(d��)��������_(d��)ˮƽͨ�^Western blot�����߽M�������ڵ���ˮƽ���M(j��n)һ���_�C����(sh��)�����@�N�����^���a(ch��n)�����^�����[�����ƺ��Ȼ���ͻ׃�����װl(f��)���������@Щ��(sh��)��(j��)������FGFR3������һ��(g��)�dz�������������������[�����ί����c(di��n)�������[������ֳ����ϵ�y(t��ng)�ڶ���(g��)�Ҋ�Đ����[��֮һ����s40%-50%�İ����[�����F(xi��n)��FGFR3����l(f��)��ͻ׃����Ƥ�[���l(f��)���ĸ���(80%)�Ƚ���(r��n)���[��������
�S���˂���(du��)FGFR3���鲻ͬ�[���ί����c(di��n)���dȤԽ��Խ�����Լ��M(j��n)���l(f��)�F(xi��n)�����^�ɼ�(x��)�����е��^�ȱ��_(d��)���҂����_ʼʹ���ɾ��wչʾ���g(sh��)�_�l(f��)�����ί�����Դ���w���ɾ��w���wչʾ��Ŀǰ��õ�һ��(g��)�_�l(f��)�����о����R���Լ��ί���;��Դ���w�ķ��������ǣ���(du��)�ڿ��w�_�l(f��)���f��F(xi��n)GFR3���ӷdz��y����ĥ����?y��n)�С����˵�FGFR3ͬԴ�Էdz���(92%)���H������Ј�(b��o)��ͨ�^ʹ��һ��(g��)��(k��)�ݷdz���(2.1*1010)���̘I(y��)��Fab��(k��)���_�l(f��)��ᘌ�(du��)һ��(g��)FGFR3���͵�FabƬ�������҂���ԇ�(y��n)�����҂�ʹ���˃ɂ�(g��)���_��scFv���w��(k��)Tomlinson I + J (MRCGeneservices, Cambridge, United Kingdom)���ɂ�(g��)��(k��)�Ď�(k��)�ݶ������1.4*108��scFvs���IgG�Լ�Fabs���и��õ��[������(r��n)�����܉�����ٱ������ͬ�r(sh��)���и��õ��خ��������@ƪ��(b��o)���У��҂��ѽ�(j��ng)�Y�x����һЩ��(du��)FGFR3a�ā���IIIc�����خ��Ե���scFv���w���@Щ���wͨ�^FACS�C�����ԺͰ����[����(x��)��ϵRT112�l(f��)������(y��ng)���������Ƽ�(x��)����ֳ�������M(j��n)һ�������ί��ĝ�����
���Ϻͷ���
��(x��)��ϵ�����������w��
RT112��HEK293���ؽM��FGFR3a��IIIc��/Fc��F(xi��n)GFR1a��IIIc��/Fc��F(xi��n)GF9��F(xi��n)GF1�Լ���Ƥ���L(zh��ng)����������FGFR3�ο�¡���w������IgG��Fc�خ��ԣ�����c-myc�ο�¡���w���ܵ������Ƅ���HRP-��c-myc���w����6His���w����M13���w��HRP-��M13�ο�¡���w��F(xi��n)ITC-�ÿ���IgG��R-��t��������IgG����IgG TrueBlot��
FGFR3 cDNA��¡�ͼ�(x��)���D(zhu��n)Ⱦ
FGFR3�İ���^(q��)�����IgG��Fc C���ںϱ��_(d��)����(g��u)�����_(d��)�d�wpcDNA3.1-FGFR3(IIIc)WT-Fc��pcDNA3.1-FGFR3(IIIc)S249C-Fc��Ȼ���D(zhu��n)ȾHEK293T��(x��)��������ӡ�E����FACS�z�y(c��)���ױ��_(d��)�ͻ�����
���ɾ��w��(k��)�Y�xFGFR3�خ���scFv���w
��scFv�ɾ��w��(k��)Tomlin-son I + J���o���ɾ��wKM13��E.coli TG1��HB2151���քe�Ϊ�(d��)���B(y��ng)�ɂ�(g��)�ɾ��w��(k��)��Ȼ��1:1����ɾ��w���ںY�x������ʾ��D1��ʾ���M(j��n)���ɾ��w��(k��)�Y�x��scFv���_(d��)����һ݆�Y�x��ʹ��1ug FGFR3����IgG���װ����װ����ɾ��w�Ⱥ���IgG����1С�r(sh��)ȥ�����Ժ�Fc�l(f��)������(y��ng)���ɾ��w���ٺ�FGFR3�����Ŀ���2С�r(sh��)��������װ�ʹ��0.1%PBSTϴ��10��(��һ݆�Y�xϴ��20��)��ʹ��100ul�ȵ���ø̎���װ���ϴÓ�Y(ji��)�ϵ��ɾ��w��ϴÓ�@�õ��ɾ��w����Goletz�������ķ����M(j��n)����һ݆�����x��
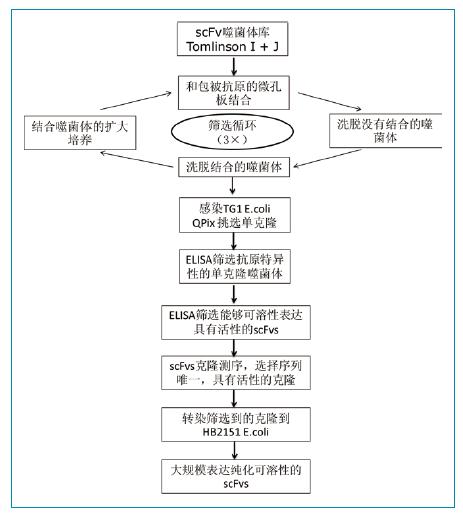
�D1���ɾ��w��(k��)�Y�xFGFR3�خ���scFv���w���̈D
�ɾ��wELISA
ʹ��0.3ug��FGFR3��F(xi��n)GFR1������IgG���װ����װ壬ϴ�������]�����벻ͬ���ǰ��Y�x��������ɾ��w�Ҹ�Һ����������HRP-��M13���w����������@ɫ��450nm�xȡ�Y(ji��)����
�ο�¡�ɾ��wELISA
��(j��ng)�^��݆����݆�ĺY�x���ի@���ɾ��w��(j��ng)�^���B(y��ng)���L(zh��ng)����¡��Ȼ��ʹ��QPix��ͨ���Ԅ�(d��ng)����¡�Y�xϵ�y(t��ng)(Molecular Devices)��ȡ�ο�¡������100ul 2TY���B(y��ng)����96�װ���37�����B(y��ng)���ڶ���ʹ����ͬ���B(y��ng)����(du��)���B(y��ng)��1:100ϡ���Ȼ��37����ʎ���B(y��ng)2С�r(sh��)������25ul����109�o���ɾ��wKM13��2TY���B(y��ng)�����^�m(x��)���B(y��ng)1С�r(sh��)�����w��(j��ng)�^�x�����ؑ���2TY���B(y��ng)����30���^ҹ���B(y��ng)��������x����ȡ50ul�������M(j��n)�Іο�¡�ɾ��wELISA������
������scFv���w�ı��_(d��)������ELISA�z�y(c��)
�@�õĸ��خ��Կ�¡��ʹ��E.coli HB2151�T��(d��o)���_(d��)���x���ի@���w����ȡ��(x��)�����|(zh��)ǻ���������ʹ���H��ɫ�V�����������ĸ��M��ʹ��SDS-PAGE�Ϳ��R˹���{(l��n)Ⱦɫ�����������ĽM���M(j��n)һ��ʹ�÷��ӺY�������x��scFv���Ć��w(scFv���װl(f��)�������w���߶���w)��
������x���w�������
ʹ��BiacoreX����������scFv��FGFR3�ĽY(ji��)�τ�(d��ng)���W(xu��)������Ӌ(j��)��@��ÿ��(g��)������scFv��Kdֵ��ʹ�ø�(j��ng)��(zh��ng)�ԽY(ji��)�Ϸ����_��ÿ��(g��)scFv���خ��ԽY(ji��)��λ�c(di��n)��ʹ��ͬ�ӵķ�������scFvs��FGF9��F(xi��n)GF1�Լ�EGF�Ƿ��ܸ�(j��ng)��(zh��ng)�Y(ji��)��FGFR3��
��ʽ��(x��)���x���۽��@�R����
ʹ����ʧ��(x��)���x�����ɾ��wscFv�ͼ����������scFv�ڼ�(x��)��ˮƽ�Ϻ�FGFR3�ĽY(ji��)�ϻ�����ͨ�^̎��RT112��(x��)����Ȼ��ʹ�ù��۽��@�R�M(j��n)һ���������w�Y(ji��)�ϻ�����
��(x��)����ֳ����
ʹ�ò�ͬ��ȵĿ�FGFR3 scFvs���w(0.02-2umol/L)̎��RT-112��(x��)����48С�r(sh��)����ʹ��MTT��(du��)��(x��)���M(j��n)��Ⱦɫ��Ȼ��570nm�x��(sh��)����(x��)������ͨ�^���湫ʽӋ(j��)�㣺Abs-scFv̎���ļ�(x��)��/Abs-��(du��)�սM��(x��)����
�Y(ji��)��
�Y�xFGFR3�خ���scFv���w�����1��ʾ�������S�C(j��)���x��288��(g��)��¡��ͨ�^ELISA�z�y(c��)���@��15��(g��)FGFR3�خ��ԵĿ��w��¡��ͨ�^�y(c��)��l(f��)�F(xi��n)��15��(g��)��¡������6��(g��)��¡��������Ψһ����3A��1D���Ѓ�(n��i)����һ��(g��)�Kֹ�ܴa����ֻ��VL��CDR2�^(q��)��һ��(g��)�A��ͻ׃������4��(g��)��¡�������б��^��ą^(q��)�e����Ҫ�^(q��)�e������CDR2��CDR3������CDR3��VH��VL���Ǹ߶ȿ�׃�ģ�CDR2��ͻ׃��Ҫ������VH��
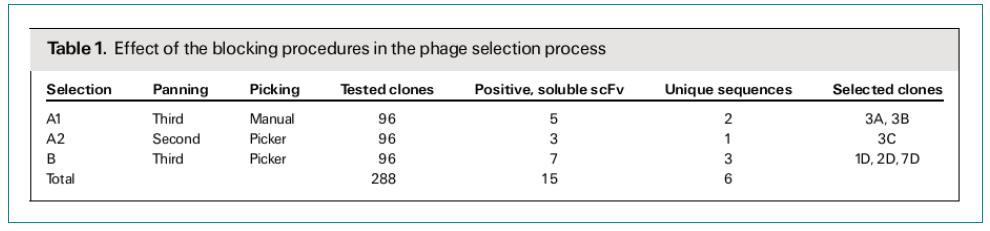
ELISA�خ����b���l(f��)�F(xi��n)����D2��ʾ���M��FGFR1��FGFR3���и߶�ͬԴ��(>62%)��6��(g��)�Y�x����scFv��¡����(du��)FGFR3�����@���ĽY(ji��)���خ������෴��FGFR1�ĽY(ji��)�ϻ��Էdz���������Ԍ�(du��)����IgG�ķ���(y��ng)�Y(ji��)�������
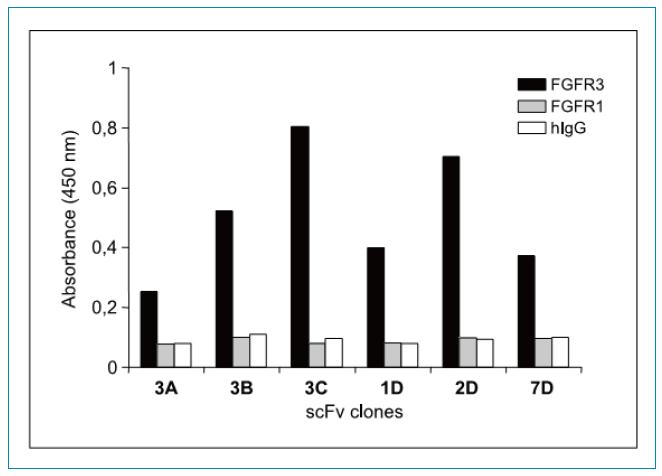
�D2��ELISA�z�y(c��)�Y�x���Ŀ�����scFvs�Y(ji��)���خ����������B(y��ng)�����Ы@�õ�scFv-pIII�ںϵ��ף�ͨ�^ELISA�����z�y(c��)��FGFR3-Fc(��ɫ)��F(xi��n)GFR1-Fc(��ɫ)�Լ���Ԍ�(du��)����IgG(��ɫ)�ĽY(ji��)�ϻ�����
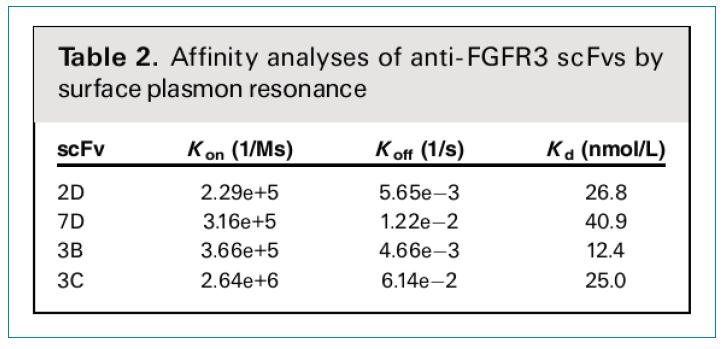
ʹ��Biacore�����˼�������wscFv���w��FGFR3���H���������2��ʾ���z�y(c��)�˲�ͬscFvs��Kon��Koff�Լ�Kdֵ��Kdֵ��40.9��12.4nmol/L֮�g׃���������Y�x����scFv�����еȵ��^�ߵ��H�ͻ�����������]��ʹ�õ��ɾ��w�Ď�(k��)��ֻ��1.6*108�������еȴ�С��(k��)�����@��(g��)�Y(ji��)���ѽ�(j��ng)�dz�������
scFv���w��(du��)RT112��(x��)�����_(d��)��FGFR3�Ļ��ԣ�ʹ��FACS�z�y(c��)��scFvs�Ͱ��װ�RT112��(x��)�����_(d��)����ȻFGFR3���ĽY(ji��)�ϻ��ԡ���D3A���������șz�y(c��)��6��(g��)scFvs�ɾ��w��¡��6��(g��)��¡�����ԺͰ��װ�RT112��(x��)���l(f��)���@���ķ���(y��ng)��Ȼ�����z�y(c��)�������4��(g��)scFvs����(3B��3C��2D��7D)�ĽY(ji��)�ϻ������l(f��)�F(xi��n)3C��7D���Ժ�RT112��(x��)���l(f��)���@���ķ���(y��ng)��2D��3B�ķ���(y��ng)��������(du��)�^����ʹ�ù��۽��@�R�^����3C��7DȾɫ�ļ�(x��)�����M(j��n)һ���@ʾ��scFv��Ĥ����Ⱦɫ����(�D3A)��
ͨ�^�ڷ�׃�ԗl���µ����߳��������M(j��n)һ���C��scFv��FGFR3���H���خ�������D3B��ʾ��������ăɂ�(g��)scFv��3C��7D���������_��λ�îa(ch��n)���@���ĵ��חl�����M(j��n)һ���C���@�ɂ�(g��)scFv���w�����R(sh��)�e�����Եĺ���Ȼ��FGFR3���w��
scFv���w��FGFR3ͻ׃�w�ķ���(y��ng)���ԣ���?y��n)�FGFR3�ڰ��װ���(x��)���н�(j��ng)���l(f��)��ͻ׃�����ԣ�Ҳ�z�y(c��)�˃ɂ�(g��)scFv���w��FGFR3 S249Cͻ׃�w�ĽY(ji��)�ϻ��������Ј�(b��o)��FGFR3 S249C���ڰ��װ���(x��)�����(j��ng)���l(f��)����ͻ׃��ʽ��ͨ�^PCR������FGFR3�����������ͻ׃����ͨ�^DNA�y(c��)���C��ͻ׃λ�c(di��n)���_(��D4A)��ͨ�^western bolt�(y��n)�C�D(zhu��n)Ⱦ��HEK293T��(x��)���������_���_(d��)ͻ׃��FGFR3S249C����(��D4B)������FGFR3S249C���к�Ұ����FGFR3WT����Ƶı��_(d��)ˮƽ��FACS�����C���ɂ�(g��)scFvs 3C��7D�������R(sh��)�e���_(d��)��ͻ׃FGFR3����HEK293T��(x��)���������ȱ��_(d��)Ұ����FGFR3���ļ�(x��)���ķ���(y��ng)���Ը���(qi��ng)(�D4C)��
����RT112��(x��)����ֳ���҂��z�y(c��)��scFvs���w������RT112�[����(x��)����ֳ����Ч�ԡ���������(g��)��ͬ��ȵ�scFv��RT112��(x��)������D5A��ʾ���ɂ�(g��)scFv 3C��7D���܉��@�����Ƽ�(x��)����ֳ����������Ч��(y��ng)�ͼ���Ŀ��w��ȳ������P(gu��n)��2umol/L��ȵĕr(sh��)�����ƻ����(qi��ng)��������0.2umol/L�ĕr(sh��)����Ȼ�����@�������ƻ�����
����֮�����z�y(c��)�˲�ͬ�����L(zh��ng)���ӌ�(du��)scFvs����Ч��(y��ng)��Ӱ�����D5B��ʾ��scFvs������Ч��(y��ng)���е��������خ�����3C��7D������������FGF9�T��(d��o)����ֳ��������ֻ��7D�܉�����FGF1�T��(d��o)����ֳ���ɂ�(g��)scFvs���w����������EGF�T��(d��o)�ļ�(x��)����ֳ����������������Ե�FGFR3�������������w��scFv������Ч�������ΑB(t��i)�W(xu��)�Ͽ���3C��7D̎��������(x��)���ΑB(t��i)��(hu��)�l(f��)��׃����������7D̎���������@��(d��o)�¼�(x��)�������γ�(�D5C)��
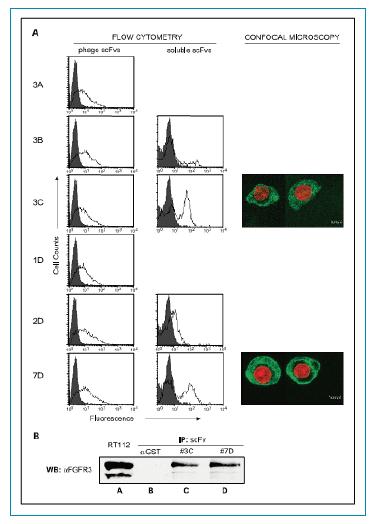
�D3.FACS���۽��@�R�����ɾ��w��������scFvs����RT112���װ���(x��)���ĽY(ji��)�ϻ�����A. FACS ֱ���D�@ʾÿ��(g��)scFv��¡�ĽY(ji��)�ϻ���(�־�)�Լ������ɹ�(��ɫ)��RT112��(x��)��ʹ��3C ��7D scFvs��(bi��o)ӛ��ʹ��FITC�ɹ⿹�w�Լ�DAPIȾɫ��(x��)��������۽��@�R�³����@ʾ(�Gɫ)scFvs���Y(ji��)�ϼ�(x��)��Ĥ��(�tɫ)DAPIȾɫ��(x��)���˅^(q��)����B. RT112��(x��)���ѽ�ҺFGFR3 IP������A.RT112��(x��)���ѽ�Һ��B. ��Ԍ�(du��)����C, scFv3C��D, scFv7D��
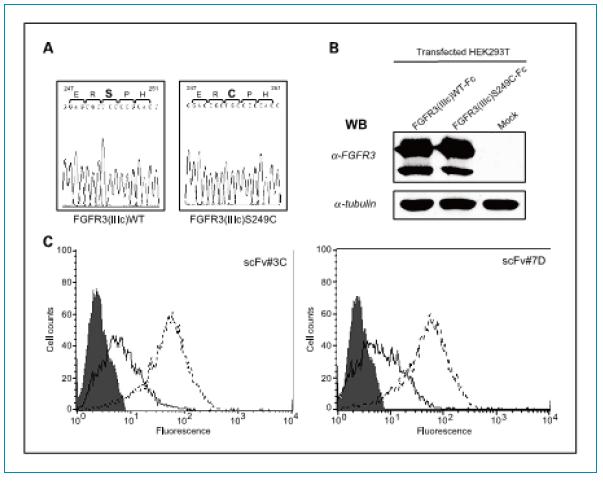
�D4.�Y�x����scFvs��FGFR3ͻ׃�w����^(q��)��ķ���(y��ng)������A.�y(c��)���@ʾDNA����������ͻ׃��Ser(TCC)249 ͻ׃?y��u)�Cys (TGC)��B. ����ӡ�E�����C��Ұ����FGFR3��ͻ׃FGFR3 (S249C) ��HEK293T��(x��)���еı��_(d��)��C . F A C S ����s c F v ���ͱ��_(d��)Ұ����FGFR3��ͻ׃FGFR3 (S249C) HEK293T��(x��)���ĽY(ji��)�ϻ���������D�У���ɫ��ʾ��Ԍ�(du��)������ɫ����ʾFGFR3(IIIc)WT ��̓����ʾFGFR3(IIIc)S249C��
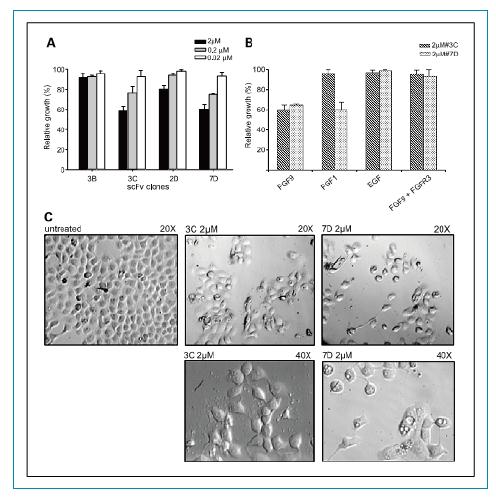
�D5. FGFR3�خ��Ե�scfvs���w��(du��)RT112��(x��)����ֳ��������A. ��ͬ��ȵ�3B, 3C, 2D��7D scFvs���w��(du��)RT112��(x��)�������ƻ��ԡ�C. 2 umol/L 3C and 7DscFvs̎����(x��)����(x��)����(sh��)Ŀ���ΑB(t��i)�ķ�����
ӑՓ
FGFR3��һ��(g��)���w�Ұ��ἤø��ͨ�^���ͬ����̖(h��o)ͨ·�ڼ�(x��)�����L(zh��ng)���[���γ��^���аl(f��)�]���������˰��װ���F(xi��n)GFR3���S������[���аl(f��)��ͻ׃���^���_(d��)������FGFR3������һ��(g��)��(y��u)�����Ч���ί����[�����c(di��n)����ǰ���о������������Ե���ȻFGFR3�İ������w�Y(ji��)�υ^(q��)��������Ƽ�(x��)����ֳ��Ч��(y��ng)��ԭ�����ǿ��Ը�(j��ng)��(zh��ng)�Y(ji��)�����w���p�����w�ͼ�(x��)���������w�Y(ji��)�������wƬ�ο���ģ����ƵęC(j��)�Ɓ����Ƽ�(x��)����ֳ����(sh��)�����S���о��ѽ�(j��ng)������3C��7D�܉�����RT112��(x��)����ֳ�ęC(j��)�����ǿ��wƬ��ֱ����ֹ��FGF-FGFR3����������Ķ�������FGFR3�ļ���������3C��7D�Y(ji��)��FGFR3��Ľ��x�ٶ�̫��(��Koffֵ̫�s
1.22*10-2��6.14*10-2)��Koff�Ĝp�٣���ζ��Kdֵ�����ӣ������w�H�������������ԣ�������3C��7D scFv�����О�ģ�����ڴ˻��A(ch��)��ͨ�^CDR�^(q��)ͻ׃�����w��(x��)��ͻ׃ģ���M(j��n)���H����������
�����Y��
1. Jorge Mart��nez-Torrecuadrada, Gabriela Cifuentes, Paula L��pez-Serra, et al., Targeting the Extracellular Domain of Fibroblast Growth Factor Receptor 3 with Human Single-Chain Fv Antibodies Inhibits Bladder Carcinoma Cell Line Proliferation. Clin Cancer Res 2005;11:6280-6290.
��(bi��o)����
Qpix 400 �ɾ��w Molecular Devices

- Odin™��(x��)�����L(zh��ng)��(d��ng)���W(xu��)�z�y(c��)�x����ȫ���˽���������L(zh��ng)��r
- �Ԅ�(d��ng)Ӌ(j��)�y(c��)�VĤ��(y��ng)�ð�����Ӌ(j��)�y(c��)�������ˮ�ӱ��д��c��Ⱥ����(sh��)��
- ȫ�Ԅ�(d��ng)���B(y��ng)����b�x��(y��ng)�ð������������ˮ���係��(sh��)�z�(y��n)
- �Ԅ�(d��ng)����Ӌ(j��)��(sh��)�x�������о��係��(sh��)�z�(y��n)�еđ�(y��ng)��
- �����Һ�w�w��Ӌ(j��)��(sh��)���������~���w���ܿ،�(sh��)�(y��n)����
- PIXL����߾������x����վ���־�Ȧ�о��еđ�(y��ng)��
- ��������z�l(f��)���������ﰲȫ�O(sh��)ʩ�I(l��ng)����о�����(y��ng)��
- QPix420�����¡�Y�xϵ�y(t��ng)�����������еđ�(y��ng)��
- ���ݴ������ʮ����Ľ����Ϻ���������չ
- ��32�óɶ��t(y��)���������[��(hu��)��Ո(q��ng)��
- �ؑc��ѩ�Ƽ���Ʒ T90��(x��)��Ϳ���ӷN�x
- ���ݴ����� MicroRisk 2024 �����L(f��ng)�U(xi��n)�u(p��ng)�����(hu��)
- Sterisart® Universal ���Ĵ��o���z�y(c��)����Ʒ����
- ��̩ؐ��Ո(q��ng)����NOVA���g(sh��)ɳ��ɳ����5.31̖(h��o)��
- �_(d��)��ȫ�Ԅ�(d��ng)�������L(zh��ng)���������x�\(ch��ng)��ȫ��(gu��)��(j��ng)�N��
- ��������ӱ�̎��ϵ�y(t��ng)AUTOPLAK���У��\(ch��ng)�д���


Copyright(C) 1998-2024 �������ľW(w��ng) �Ԓ��021-64166852;13621656896 E-mail��info@bio-equip.com